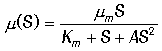|
(14)
|
Микробиологические популяции являются хорошим экспериментальным объектом для
проверки идей и результатов как экологических, так и эволюционных теорий. В
большинстве своем микроорганизмы - одноклеточные организмы, они имеют высокое
отношение поверхности к объему и поэтому высокие интенсивности обмена с
окружающей средой, высокие скорости размножения, большой прирост биомассы. Для
математического описания микробных популяций обычно используют аппарат
обыкновенных дифференциальных уравнений. В отношении микробиологических систем
такое описание гораздо более обосновано, чем применительно к наземным и водным
высшим организмам. В лабораторных исследованиях, in vitro работают с количеством
особей порядка 1010 и выше. В большом
промышленном ферментере могут одновременно жить 1016
- 1017 дрожжевых клеток. Отклонение численности от средних
значений, вызванное случайными обстоятельствами, пропорционально 1/N1/2, где N - численность
популяции. Таким образом, для многочисленных популяций можно строить модель в
терминах средних численностей, или концентраций. Второй фактор - относительная
однородность культуры микроорганизмов в объеме культиватора. Это позволяет
пренебречь пространственными эффектами.
В микробиологии общепринят эмпирический подход к построению моделей. Из всех факторов, влияющих на рост клетки, выбирают лимитирующий, и опытным путем находят зависимость скорости роста от его концентрации. В общем виде кинетика концентрации клеток в непрерывной культуре описывается уравнением:
dx / dt = x( |
(13)
|
Здесь x – концентрация клеток в культиваторе; mx, концентрации субстрата (обычно обозначается S), температуры, рН среды и прочих
факторов; n
Для поддержания культуры в области нелимитированного роста требуются внешние
регуляторы. В случае лимитирования роста внешним фактором, например, недостатком
субстрата, стационарный режим работы культиватора устанавливается путем
саморегуляции. Это имеет место в природных проточных системах и в наиболее
распространенном типе непрерывных культиваторов - хемостате, где задается скорость разбавления культуры, или
скорость протока. Теория хемостата - впервые была разработана Моно (1950) и Гербертом (1956) и с той поры постоянно
совершенствуется. В современных моделях учитывается структурная неоднородность
биомассы, возрастная
неоднородность культуры и другие детали культивирования.
При непрерывном перемешивании можно считать весь объем культиватора однородно заполненным, концентрации субстрата и клеток в каждой точке культиватора одинаковыми, и описывать поведение этих концентраций во времени с помощью системы обыкновенных дифференциальных уравнений:
|
(14)
|
Здесь S - концентрация субстрата; x - концентрация клеток в культиваторе; S0 -концентрация субстрата, поступившего в
культиватор; D - скорость протока (разбавления) культуры;
a-1m(S)x-Dx - отток
биомассы из культиватора; - - количество субстрата,
поглощенного клетками культуры; am(S)xDS0 - приток
субстрата в культиватор; -DS - отток неиспользованного
субстрата из культиватора. Скорость роста биомассы предполагается зависящей
только от концентрации субстрата в соответствии с формулой Моно (5).
Рассмотренная модель является упрощенной и для описания реальных процессов требует дополнений. Например, при больших концентрациях субстрат может оказывать ингибирующее действие, и тогда формулу для скорости роста следует записывать в виде:
|
(15)
|
В системе , где существует такая зависимость скорости роста от субстрата,
возможны триггерные
режимы - наличие двух устойчивых стационарных
состояний и зависимость стационарных значений концентраций субстрата и
биомассы от начальных условий (от величины затравки и начальной
концентрации биомассы).
На скорость роста биомассы может оказывать влияние концентрация продуктов метаболизма в среде, окружающей клетку. Тогда к двум уравнениям, описывающим динамику концентрации биомассы и субстрата в непрерывном процессе культивирования следует добавить третье уравнение, выражающее динамику концентрации продуктов метаболизма
|
(16)
|
Формула (16)
известна как формула Моно-Иерусалимского [.
В биотехнологии для расчета оптимальных режимов культивирования применяются формулы, принимающие в расчет и другие особенности как метаболизма самих микроорганизмов, так и условий их выращивания.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ