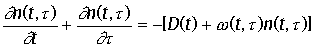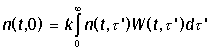Непрерывные модели возрастной структуры
Непрерывные модели оперируют не с численностями отдельных групп, а с
непрерывной функцией распределения организмов по возрастам. Уравнение для
плотности функции распределения было впервые получено Мак-Кендриком в 1926 г., а затем "переоткрыто" фон Ферстером в 1959 г. и носит его имя. Это уравнение
представляет собой дифференциальную форму закона сохранения числа особей. В
уравнении две независимые переменные - время t и возраст
t, который отсчитывается с
момента рождения особи. n(t, t) dt - количество особей, имеющих возраст
в интервале [t,t+dt]. Общее число особей всех возрастов
в момент времени t определяется интегралом 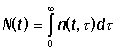 . Уравнение Ферстера имеет вид:
. Уравнение Ферстера имеет вид:
с начальным условием n(0, t) = g(t)
В уравнении (24)
слева стоит полная производная dn / dt, при этом учтено, что dt / dt = 1, в правой
части уравнения - члены, которые описывают процессы, приводящие к изменению
числа клеток данного возраста. Убыль клеток может быть вызвана разными причинами
- смертностью, миграцией, для проточной культуры всеми этими процессами можно
пренебречь по сравнению с протоком клеток через культиватор. Скорость протока
D(t) не зависит от возраста клеток, но может зависеть от
времени. Член -w(t,
t) u(t, t) описывает убыль клеток из данного
интервала возрастов при делении на дочерние со скоростью w. Прирост численности в результате
размножения происходит в нулевой возраст и войдет в граничное условие при t = 0
Здесь k - число потомков в одном акте размножения,
W(t, t') dt' - вероятность размножения родителя
в возрастном интервале [t', t' + dt'], равная удельной скорости
размножения;
Если родители остаются в популяции после размножения (дрожжи), то W(t, t) - плотность
безусловной вероятности деления в возрасте t (функция распределения возрастов деления).
Если же клетки выбывают из своей возрастной группы после деления (водоросли,
бактерии), то W(t, t) - плотность условной вероятности
разделиться в возрасте t,
если клетка дожила до этого возраста, не разделившись.
Имеются модели, описывающие распределение клеток по размерам и массам. Их
легче сопоставлять с экспериментальными данными, так как имеются
экспериментальные методы определения размеров клеток. Активно разрабатываются
методы микроизмерений, позволяющие определить и другие параметры отдельных
клеток (например, фотосинтетическую активность, содержание хлорофилла в
водорослях, внутриклеточное рН и др.). Все большее распространение получают
методы проточной микрофлуорометрии, позволяющие регистрировать спектральные
характеристики сотен и тысяч микроорганизмов и строить соответствующие
распределения признаков отдельных особей. Информация об эволюции этих
распределений дает новые возможности оценки состояния популяций микроорганизмов,
например, состояний популяций планктона в морях, почвенных микроорганизмов,
клеток крови.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ