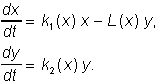 (7.8)
(7.8)
С середины 20 века в связи с развитием интереса к экологии и с быстрым усовершенствованием компьютеров, позволившим численно решать и исследовать системы нелинейных уравнений, стало развиваться направление популяционной динамики, посвященное выработке общих критериев с целью установить, какого вида модели могут описать те или иные особенности поведения численности взаимодействующих популяций, в частности, устойчивые колебания.
Эти работы развивались по двум направлениям. Представители первого направления, описывая входящие в модельные системы функции, задают лишь качественные особенности этих функций, такие как положительность, монотонность отношения типа большеменьше (Колмогоров, 1972, Rosenzweig, 1969). Рассматриваемые здесь модели могут быть изучены аналитически.
В рамках второго направления последовательно рассматривались различные модификации системы Вольтерра, получаемые включением в исходную систему различных дополнительных факторов и закономерностей, описываемых явными функциями (Холлинз, 1965, Иевлев,1955, Полуэктов, 1980, Базыкин, 1985, Медвинский, 1995). Использование компьютерной техники позволило применить полученные здесь результаты к конкретным популяциям, в частности, к задачам оптимального промысла.
Примером работ первого направления служит работа А.Н.Колмогорова (1935, переработана в 1972), который рассмотрел обобщенную модель взаимодействия биологических видов типа хищникжертва или паразит-хозяин. Модель представляет собой систему двух уравнений общего вида
(7.8)
В модель заложены следующие предположения:
1) Хищники не взаимодействуют друг с другом, т.е. коэффициент размножения хищников k2 и число жертв L, истребляемых в единицу времени одним хищником, не зависит от y. 2) Прирост числа жертв при наличии хищников равен приросту в отсутствие хищников минус число жертв, истребляемых хищниками. Функции k1(x), k2(x), L(x), - непрерывны и определены на положительной полуоси x,y 0. 3) dk1/dx<0. Это означает, что коэффициент размножения жертв в отсутствие хищника монотонно убывает с возрастанием численности жертв, что отражает ограниченость пищевых и иных ресурсов. 4) dk2/dx>0, k2(0)<0<k2(). С ростом численности жертв коэффициент размножения хищников монотонно убывает с возрастанием численности жертв, переходя от отрицательных значений, (когда нечего есть) к положительным. 5) Число жертв, истребляемых одним хищником в единицу времени L(x)>0 при N>0; L(0)=0.
Исследование этой модели и ее частных случаев, например, модели Розенцвейга (1965,1969), привело к выводу о том, что регулярные колебания в системе имеют место, если численность хищника ограничивается наличием жертвы. Если численность жертвы ограничивается количеством необходимых ей ресурсов, или численность хищника ограничивается не количеством жертвы, а другим фактором, это приводит к затухающим колебаниям. К затуханию колебаний приводит также наличие убежищ для жертв, которые делают их недоступными для хищников. Амплитуда колебаний будет возрастать, и это приведет в конце концов к вымиранию одного или обоих видов, если хищник может прокормиться при такой плотности популяции жертв, которая значительно ниже допустимой емкости среды (которая следует из логистического уравнения.
Модель взаимодействия двух видов насекомых (MacArthur, 1971) является одной из наиболее известных моделей, которая использовалась для решения практической задачи - борьбы с вредными насекомыми с помощью стерилизации самцов одного из видов. Исходя из биологических особенностей взаимодействия видов была написана следующая модель
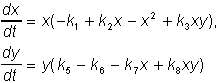 (22)
(22)
Здесь x,y - биомассы двух видов насекомых. Насекомые вида х поедают личинок вида y (член k3y), но взрослые особи вида y поедают личинок вида х при условии высокой численности одного или обоих видов. При малых x смертность вида x выше, чем его собственный прирост. Последний член во втором уравнении отражает прирост биомассы вида y за счет поедания взрослыми насекомыми вида y личинок вида x. Фазовый портрет системы изображен на рис. 10.
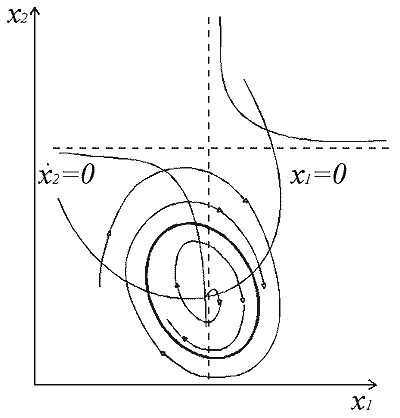 Рис.10. Колебания численностей в системе двух видов насекомых Фазовый портрет включает предельный цикл - замкнутую изолированную фазовую траекторию, описывающие автоколебания в системе. (MacArthur, 1971) |
Введение в систему управления, направленного на уничтожение вредного вида возможно в двух формах. Управление может сводиться к кратковременному, скачкообразному уничтожению одного или обоих видов, например, с помощью химических инсектицидов. Из модели видно, что для популяции из двух видов такой метод борьбы будет малоэффективным, так как с течением времени численности видов опять возрастут, и система снова выйдет на предельный цикл, то есть численности насекомых будут совершать регулярные колебания.
Второй метод - изменить характер функций, стоящих в правых частях уравнений и определяющий характер размножения и взаимодействия видов. Именно такому, параметрическому, способу отвечают биологические методы борьбы. Интересно, что в некоторых случаях оказывается, что воздействие более целесообразно приложить не к самому вредителю, а к его конкуренту. Именно такой способ был применен на практике - параметры одного из видов были изменены путем добавления стерилизованных самцов, и систему удалось привести к стационарному состоянию с малой численностью обоих видов.
Теоретический анализ моделей взаимодействий видов в книге А.Д.Базыкина "Биофизика взаимодействующих популяций" (М., Наука, 1985), а также в монографиях: Р.А.Полуэктов, Ю.А.Пых, И.А.Швытов Динамические модели экологических систем", Б.Г.Заславский Р.А.Полуэктов "Управление экологическими системами" (1988).
Особый интерес для практики представляет выработка критериев близости системы к опасным границам, при переходе через которые система перестает существовать или переходит в качественно иное состояние. При этом характер динамики популяции резко меняется, например, популяция переходит от монотонного роста к резким колебаниям численности, или просто вымирает. Такие границы называются бифуркационными. Исследование свойств моделей показывает, что одним из признаком близости к опасной границе является очень медленное восстановление численности после воздействия неблагоприятного фактора. Индикатором опасности служит также изменение формы колебаний численностей хищника и жертвы. Если из близких к гармоническим колебания становятся релаксационными, то есть характерные времена изменения численности видов начинают сильно различаться, причем амплитуда колебаний со временем нарастает, это может привести к потере устойчивости системы и вымиранию одного или обоих видов.
Мы рассмотрели простейшие модели популяционной динамики: модели одного или двух взаимодействующих видов. Такие модели допускают аналитическое исследование, и потому достаточно полно изучены. Дальнейшее углубление математической теории идет по линии детализации структуры самих популяций (возрастной, половой) и учета временных и пространственных факторов.
Учет запаздывания в моделях взаимодействующих видов приводит к системам уравнений, изучение существования и устойчивости которых представляет серьезную математическую проблему, а сами решения, описывающие динамику численности, могут носить не только колебательный, но и квазистохастический характер, как это мы видели для дискретных моделей популяций с неперекрывающимися поколениями. Устойчивость или неустойчивость модели может зависеть от периода запаздывания, и зависимость эта может быть немонотонной и даже многоэкстремальной. В последние годы для систем с запаздыванием используется понятие абсолютной устойчивости, справедливое для систем, которые локально устойчивы независимо от величины запаздывания.
Обычно не все члены уравнений содержат запаздывание. Например, увеличение численности популяции может сказаться на скорости роста лишь через определенное время, когда молодые особи достигнут репродуктивного возраста, а хищничество сокращает численность жертв тот же момент времени и не содержит запаздывания. Исследование учитывающих эти обстоятельства моделей показывает, что саморегулирование жертв стабилизирует систему, а запаздывание в реакции хищника - дестабилизирует. Короткие времена запаздывания в размножении как хищника так и жертвы могут как стабилизировать, так и дестабилизировать систему. Последний вывод опровергает общепринятое ранее представление о том, что малое запаздывание слабо сказывается на поведении системы. Интуитивное представление о том, что чем больше запаздывание. тем больше его дестабилизирующий эффект, неправильно. В некоторых системах взаимодействующих видов оказалось, что малые времена запаздывания наиболее опасны для стабильности системы.
Моделирование конкретных сообществ часто требует подробной детализации половой и возрастной структуры. Взаимодействие возрастных групп даже в пределах одного вида приводит к сложным типам динамики численности. Рассмотрим в качестве примера модель, описывающую динамику численности насекомых с полным метаморфозом с двумя активными стадиями имаго и личинок. Уравнения имеют вид:
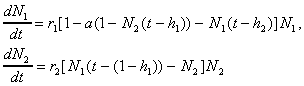 (23)
(23)
Здесь 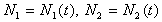 - нормированные количества имаго и личинок, h1 - время между появлением личинок и имаго, (1- h1) - время между появлением имаго и личинок, r2 - скорость появления личинок, 0<a<1, r(1-a)=r1 - мальтузианский коэффициент линейного роста, который определяется по плодовитости имаго, h2 - среднее время жизни в течение одного года популяции имаго. На рис. 10 показана динамика численности переменных в модели: (а) - колебания численностей, (б) - реализацию случайного процесса с большими всплесками численности и отдельными периодами почти полного исчезновения особей.
- нормированные количества имаго и личинок, h1 - время между появлением личинок и имаго, (1- h1) - время между появлением имаго и личинок, r2 - скорость появления личинок, 0<a<1, r(1-a)=r1 - мальтузианский коэффициент линейного роста, который определяется по плодовитости имаго, h2 - среднее время жизни в течение одного года популяции имаго. На рис. 10 показана динамика численности переменных в модели: (а) - колебания численностей, (б) - реализацию случайного процесса с большими всплесками численности и отдельными периодами почти полного исчезновения особей.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ