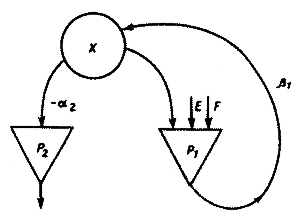
 Рис.4. Схема растения как системы с лимитирующими факторами. |
В силу сложности процессов в экологической системе необходимо выделить главные факторы, взаимодействие которых качественно определяет судьбу системы. Фактически все модели, включающие описание роста популяций или сообществ, основываются либо на "принципе лимитирующих факторов" (Leibig,1840. Перевод на русский язык: Либих, 1936), либо на "законе совокупного действия факторов", Э.А.Митчерлихf (Mitscherlich, 1909, 1925). Исходно эти принципы были сформулированы для популяций одного вида, однако применяются для описания многовидовых сообществ и экосистем.
Концепция лимитирующих факторов принадлежит немецкому агрохимику Юстусу Либиху , который предложил знаменитый закон минимума:. "Каждое поле содержит одно или несколько питательных веществ в минимуме и одно или несколько других в максимуме. Урожаи находятся в соответствии с этим минимумом питательных веществ".Либих понимал под этим относительный минимум питательного вещества по сравнению с содержанием других веществ. Позже в экологической литературе фактор, находящийся в минимуме, стали называть лимитирующим фактором. Закон "лимитирующего фактора" для фотосинтетических процессов в 1905 г. предложил Ф.Блэкман, а в 1965 г. Н.Д.Иерусалимский сформулировал этот закон для ферментативных процессов. Естественно, что при изменении соотношений факторов, лимитирующий фактор может изменяться.
Математическая теория описания систем с лимитирующими факторами разработана И.А.Полетаевым и его школой (Полетаев, 1966, Гильдерман, 1970). Обобщение принципа лимитирующего фактора с использованием теоретико-множественных представлений и разработка методов выделения лимитирующего звена с энтропийных экстремальных принципов дана в работах А.П.Левича (1988, 1994). Разработан класс дискретно непрерывных моделей, описываемых системами дифференциальных уравнений, структура которых меняется в определенные моменты времени, а последние в свою очередь зависят от значения решений самих уравнений. И.А. Полетаевым эти системы названы системами с лимитирующим фактором, или Л-системами.
Самым простым примером Л-системы служит химическая реакция соединения нескольких веществ в одно, происходящая практически мгновенно. Вещества-реагенты могут взаимодействовать лишь в строго определенных стехиометрических соотношениях, и поэтому скорость рассматриваемой реакции определяется самой медленной в данный момент времени из скоростей притока каждого из исходных веществ в сферу реакции. Иными словами, свойства Л-системы определяются наличием узкого места и его сменой в процессе функционирования.
Метод описания элемента экологической модели как Л-системы можно проиллюстрировать на примере простой модели роста растения (Полетаев, 1973). Основные процессы превращения вещества и энергии в растениях, это - процесс возрастания биомассы (роста), идущий за счет фотосинтеза и поглощения веществ из почвы и атмосферы, и процесс основного обмена (дыхания), поставляющий свободную энергию для жизнедеятельности биомассы за счет частичного расходования вещества, содержащегося в самой биомассе. Блок-схема растения изображена на рис. 4. На
Рис.4. Схема растения как системы с лимитирующими факторами. |
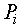 потребляет извне свет Е и некоторое вещество F, дает прирост биомассы x; процесс дыхания
потребляет извне свет Е и некоторое вещество F, дает прирост биомассы x; процесс дыхания 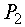 потребляет часть биомассы, поставляя свободную энергию для жизнедеятельности оставшейся части, а выделяет продукты, которые в данной модели не учитываются. Процесс роста биомассы растения описывается уравнением:
потребляет часть биомассы, поставляя свободную энергию для жизнедеятельности оставшейся части, а выделяет продукты, которые в данной модели не учитываются. Процесс роста биомассы растения описывается уравнением:
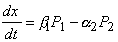 (3)
(3)
x(t) - значение биомассы в момент времени t. По смыслу задачи эта функция непрерывна и имеет производную. Пусть 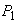 и
и 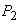 -две действительные неотрицательные переменные - интенсивности роста и дыхания.
-две действительные неотрицательные переменные - интенсивности роста и дыхания.
Величина интенсивности процессов Pj в момент времени t определяется в Л-системе лимитирующей из всех входных компонент, т.е. той, которая обеспечивает наименьшую интенсивность Pj. Все остальные компоненты, потоки которых превышают минимальный, оказываются в избытке и потребляются лишь частично. Лимитирующий фактор j-го процесса меняется по величине со временем вместе с соответствующей ему интенсивностью Pj. Вследствие изменения со временем состояния системы компонента 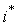 лимитирующая процесс в момент
лимитирующая процесс в момент 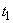 , может с течением времени перестать ею быть. Тогда начиная с момента
, может с течением времени перестать ею быть. Тогда начиная с момента 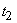 лимитирующей станет уже другая компонента
лимитирующей станет уже другая компонента 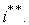 В модели растения интенсивности процессов
В модели растения интенсивности процессов 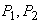 запишутся в виде
запишутся в виде
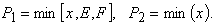 (4)
(4)
Величина потока компоненты i на вход процесса j в момент t зависит от имеющегося в системе количества этой компоненты х, а для поступающих извне компонент (Е, F) - от состояния среды. Поток внешней компоненты (вода, растворенные вещества) можно считать постоянным, пропорциональным наличию этой компоненты во внешней среде. Поток света Е предполагается пропорциональным произведению интенсивности света Ец на величину о- поглощающей поверхности растения. Если растение в процессе роста не меняет формы, т.е. сохраняет геометрическое подобие, то величину поверхности можно считать пропорциональной квадрату линейных размеров или квадрату корня кубического из величины биомассы (биомасса пропорциональна объему растений, или кубу линейного размера). Таким образом, величина потока света запишется в виде
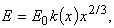 (5)
(5)
где k(x) - коэффициент, отражающий форму растения.
В зависимости от соотношений величины и параметров системы 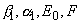 величина
величина 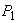 может быть равна наименьшему из трех значений. Следовательно, решение распадается на три области, в каждой из которой рост лимитируется одним из факторов.
может быть равна наименьшему из трех значений. Следовательно, решение распадается на три области, в каждой из которой рост лимитируется одним из факторов.
В последующих работах по моделированию конкретных экологических систем аппарат Л-систем нашел широкое применение. Рассмотрение биогеоценозов как систем с лимитирующими факторами позволяет "автоматически" следить за лимитированием каждого процесса. Особенно широкое применение нашел этот способ для описания микробных сообществ, где типы взаимодействия видов могут быть описаны сравнительно просто при помощи обыкновенных дифференциальных уравнений (разных, в различных областях лимитирования).
Микробные сообщества являются классическими модельными экспериментальными системами для изучения экосистемных взаимодействий. Теоретический анализ видовой структуры трофического уровня одноклеточных автотрофных и гетеротрофных организмов с точки зрения Л-систем показал, что условием сосуществования конкурирующих видов является различие определяющих факторов развития, что является подтверждением принципа Гаузе о существовании лишь одного вида в каждой экологической нише. Это условие позволяет каждому виду зависеть в большей степени от одного определенного фактора среды и в меньшей степени - от других. В случае конкуренции только за взаимозаменяемые элементы питания каждый вид имеет вообще один лимитирующий трофический фактор, отличающийся от лимитирующих факторов других видов. Эти свойства поликультур микроорганизмов учитывют при совместном культивировании нескольких видов, которое дает возможность путем изменения процентного содержания видов в сообществе регулировать химический состав суммарной биомассы и ассимиляционный коэффициент поликультуры в целом.
Л-системы являются эффективным аппаратом для изучения популяций в экологическом окружении с ограничениями, накладываемыми на их развитие наличием корма, местообитания и прочим. Устойчивые состояния и кинетика переходных процессов определяются при этом для популяции каждого вида и каждый момент времени конечным, и притом достаточно малым, числом критических ограничений для компонент узкого места.
В различных фазах жизненного цикла и в различные сезоны давление критических условий не будет одинаковым. Опознавание критических компонент и ограничений в каждом отдельном случае и их количественное изучение являются задачей экспериментальной биологии. Применение формализованного принципа лимитирующих факторов позволяет при этом формализовать классификацию явлений в биогеоценозе и более эффективно использовать результаты количественных измерений.
Концепция лимитирующих факторов оказалась весьма плодотворной в инженерной экологии (biomanipulation). Управляющими для биогеоценоза могут быть потоки только тех ресурсов, которые лимитируют рост сообщества. Поэтому разработка концепции и методов как экспериментального выявления лимитирующих факторов для многовидовых сообществ и адекватных модельных подходов представляет собой весьма актуальную практическую задачу (А.П.Левич, В.Н.Максимов, Н.Г.Булгаков. Теоретическая и экспериментальная экология фитопланктона. Управление структурой и функциями сообществ.1997)
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ